Le modèle GO+ est un modèle de croissance, de production et de gestion forestière représentant les principaux processus biophysiques et biogéochimiques d'une forêt gérée. Son approche de modélisation est de type mécaniste : elle représente les processus impliqués dans le fonctionnement de l’écosystème et les opérations de sylviculture.
Le modèle GO+ est décrit de façon détaillée dans l’article de Moreaux et al. (2020) qui sera disponible en open access. Son code informatique (Python 3) sera téléchargeable prochainement.
Le modèle considère une unité spatiale correspondant à une zone homogène de végétation forestière, typiquement un hectare, et décrit son fonctionnement et le dynamique de croissance du peuplement d‘arbres(régénération, croissance et mortalité des arbres individuels) pour l’ensemble du cycle de vie de l’écosystème forestier (50-300 ans). Le modèle décrit les échanges de masse (Carbone, Eau) et d'énergie (rayonnement, chaleur, vent) dans le système sol-végétation-atmosphère. Ces échanges déterminent le bilan d’énergie, les cycles du carbone et de l'eau. Il prend en compte les processus de transfert turbulent, transfert radiatif, l'évapo-transpiration, la photosynthèse, la respiration, la répartition du carbone dans les plantes, la croissance, la phénologie, la mortalité et les transformations et minéralisation du carbone dans le sol.
La végétation est décrite en deux couches ou strates, les arbres et la végétation accompagnatrice. Les deux couches de végétations ont dynamiques : les parties aériennes et souterraines des plantes suivent un cycle phénologique incluant les phases de dormance, débourrement, croissance et mortalité. Ce développement est rythmé par l’évolution de la température et du rayonnement solaire. La mortalité est déclenchée par les stress hydriques ou thermiques, les opérations sylvicoles et les récoltes.
Les effets sur l’écosystème des principales opérations incluses dans un itinéraire technique sylvicole donné sont pris en compte : préparation du sol, gestion du sous-étage, éclaircies, recépage, coupes à blanc.
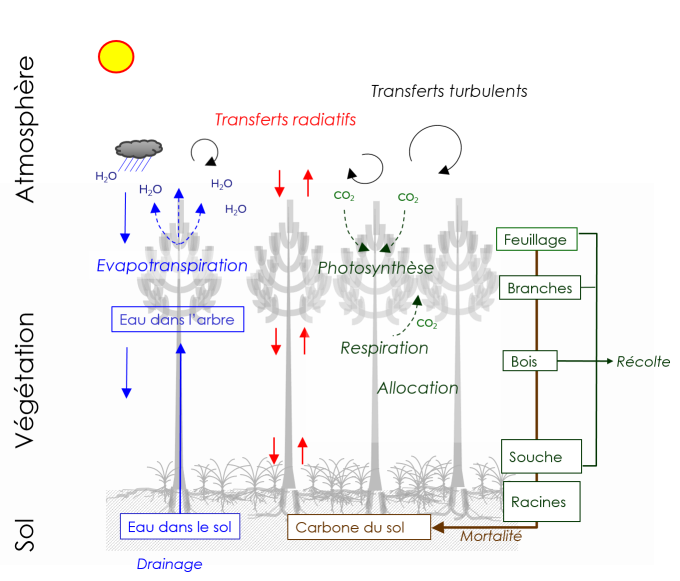
1. Transferts radiatifs et bilan d'énergie.
Le microclimat et les flux radiatifs (interception, transmission, émission) des grandes et courtes longueurs d'onde sont évalués en fonction d'un schéma simple de transfert radiatif basé sur la loi de Beer-Lambert et séparant les fractions ombrée et ensoleillée du feuillage pour chacune des couches (dePury et Farquhar, 1997).Ce schéma tient compte des fractions directes et diffuses du rayonnement solaire incident (Spitters et al. 1986) et de la rediffusion du rayonnement au premier ordre. Le bilan d’énergie et le calcul de la température du feuillage sont résolus au pas de temps horaire et pour chaque compartiment.A partir du bilan d’énergie, la température moyenne de chaque couche est calculée au pas horaire (Jones, 1992).
2. Évapotranspiration.
L’évapotranspiration est le flux de vapeur d’eau transféré depuis l’écosystème vers l’atmosphère.Ce flux représente plusieurs dizaines de mètres cubes d’eau par hectare de forêt annuellement. Il est exprimé en kg d’eau évaporé par unité de surface (m²) et de temps (heure). Un kilogramme d’eau recouvrant une épaisseur de 1 mm ur une surface de 1 m, il est couramment aussi exprimé en mm par heure (évapotranspiration). Il inclut deux composantes.
- L’évaporation depuis les surfaces mouilléeset le sol. Durant un épisode de précipitations, une fraction des précipitations est retenue par le feuillage, les branches et le tronc des arbres (interception). Une partie de l’eau ainsi interceptée estré-évaporée. Dans GO+, la quantité d’eau liquide retenue sur le feuillage et à la surface du sol durant les épisodes pluvieux est calculée au pas horaire à partir du bilan hydrique de la surface de feuillage et des éléments ligneux (troncs, branches) (Gash et al. 1979). L’autre source d’évaporation directe est le sol.
- La transpiration, depuis le couvert sec, correspond à un transport de vapeur depuis l’intérieur des feuilles vers l’atmosphère et s’opère à travers les pores stomatiques pour l’essentiel. La transpiration végétale est régulée parl’indice foliaire et le fonctionnement des stomates qui régulent la diffusion de la vapeur d’eau et au CO2. Celle-ci est modélisée en tenant compte des différents facteurs contrôlant l’indice foliaire et l’ouverture des stomates :
* Rayonnement solaire incident, température et humidité de l’air.
* Concentration en CO2 de l’air.
* Etat hydrique des feuilles (potentiel hydrique foliaire), qui dépend de la hauteur des arbres et de la disponibilité de l’eau dans le sol.
- Le transport des molécules d’eau depuis chacune des trois couches, arbres, sous-étage et sol vers l’atmosphère est décrit suivant un profil de turbulence logarithmique avec une résistance aérodynamique tenant compte de la densité des tiges, de leur hauteur et de leur indice foliaire (Nakai et al. 2008).
3. Eau dans l’arbre.
L'état hydrique du feuillage est calculé en utilisant un schéma de transfert hydraulique simple reliant l’eau du sol aux feuilles par un circuit hydraulique comprenant deux résistances en série, sol – racine et racine- feuille, et une capacitance. Le potentiel hydrique moyen du feuillage est ainsi calculé au pas horaire. Il définit l’étant énergétique de l’eau dans la plante et exprime l’état de stress hydrique de la plante. Le potentiel hydrique exerce un contrôle étroit sur l’ouverture des stomates des feuilles, et, par suite, la transpiration des arbres, la température du feuillage, la photosynthèse et la respiration du feuillage.
4. Eau dans le sol.
Le sol est décrit comme une cascade de trois réservoirs correspondant respectivement à :
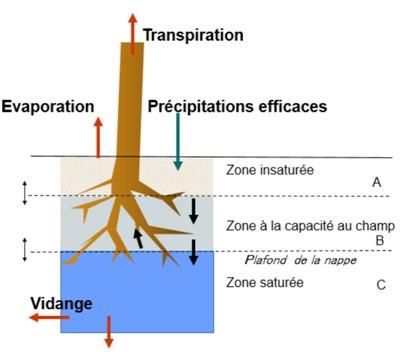
- A, zone superficielle sous la capacité au champ (beige)
- B une zone à la capacité au champ (bleu pâle)
- C une zone saturée (nappe phréatique) (bleu soutenu).
Les notions de capacité au champs, saturation et de réserve utile sont détaillées à https://appgeodb.nancy.inra.fr/biljou/fr/fiche/reserve-en-eau-du-sol. Les précipitations efficaces alimentent la couche superficielle et, par écoulement préférentiel rapide, la zone saturée. L’épaisseur des trois couches varie.Elle est calculée dans GO+ en fonction du prélèvement par les racines, du drainage profond et du ruissellement, et de la pluie efficace. La profondeur prospectée par les racines étant fixe, celles-ci peuvent solliciter différents réservoirs suivant les saisons : la nappe phréatique en période de crue comme cela est représenté dans le schéma ci-dessus, ou seulement la zone superficielle en période d’étiage. Le prélèvement racinaire sollicite celui des réservoirs prospectés par les racines où l’eau est la plus disponible, zone saturée > zone à la capacité au champ > zone sous la capacité au champ.
5. Photosynthèse et respiration des plantes.
Le calcul de la photosynthèse,ou production primaire brute, des deux couches de végétation est basé sur le modèle biochimique de Farquhar et al. (1980) le plus couramment utilisé. Partant de la cinétique enzymatique des réactions du cycle de Calvin et du transfert d’énergie réductrice créée par transformation de l’énergie lumineuse, il décrit la vitesse de fixation du carbone par le feuillage. La photosynthèse est donc exprimée en grammes de carbone-CO2 absorbé par unité de surface de sol et par unité de temps, gC m-2 jour-1. Ce formalisme, repris dans le code GO+ pour chacune des deux couches de végétation,prend en compte de façon réaliste les effets de la température du feuillage, de la concentration en CO2 et du contrôle stomatique ainsi que du rayonnement incident. La respiration de la plante est estimée pour chaque partie de la plante comme la somme de deux termes.
- la respiration de maintenance, dépendante de la température, et proportionnelle à la concentration en azote de la biomasse vivante (Ryan, 1992, Dufrêne et al. 2005). La respiration foliaire est partiellement inhibée par le rayonnement solaire.
- la respiration de croissance calculée comme le coût de construction des constituants biochimiques de la plante. Ici, ce coût est fixé à 28% du carbone utilisé pour la croissance.
Elle est exprimée comme la photosynthèse en grammes de carbone-CO2 émis par unité de surface de sol et par unité de temps.
6. Phénologie.
Pour les arbres, seul le feuillage présente un cycle annuel phénologique incluant débourrement, croissance et mortalité. La croissance des autres parties de la plante n’est résolue qu’au pas de temps annuel. Selon l’essence considérée, plusieurs modèles de phénologie foliaire sont actionnables : une somme de degrés-jour (Desprez-Loustau et Dupuis, 1994), le modèle séquentiel de Kramer (1994), ou le modèle alterné de Harrington (2010) qui sont bien adaptés au Pins, Hêtre et Douglas respectivement. La calibration des paramètres pour les essences métropolitaines s’est appuyée sur la littérature disponible et d’observations provenant du réseau Renecofor et de réseaux phénologiques aux Pays-Bas et en Allemagne.
Pour la végétation accompagnatrice, la croissance de chacune des parties de la plante, feuille, organes pérennes ou racines, est résolue au pas journalier. La phénologie répond à un modèle simple d’accumulation de degrés jours pour le démarrage de la croissance. La croissance est modélisée selon une loi de croissance potentielle sigmoïde modulée par le carbone disponible. Elle est ralentie par les stress hydrique et thermique. La mortalité des parties caduques en fin de saison est déclenchée soit par une accumulation de stress hydrique ou thermique soit par une durée limite de végétation.
7. Allocation du carbone et croissance.
L'attribution de la production primaire en carbone (GPP pour Gross Primary Production) à chaque arbre se fait au pro rata de sa contribution à l’indice foliaire. Le carbone disponible pour la croissance est réparti entre biomasses aérienne et souterraine de l’arbre selon un indice de stress (Landsberg et Waring, 1997) calculé comme le rapport entre la transpiration annuelle effective et la transpiration annuelle potentielle : plus le stress est fort plus l’allocation aux racines est élevée.
La biomasseaérienne est distribuée entre tronc, feuillage et branches selon les relations allométriques connues pour chaque espèce (Gholz, 1979, Shaiek et al. 2011, Wutzleret al. 2007, Le Moguedec et Dhôte, 2012, Achat et al. 2018). La biomasse souterraine est de même distribuée entre les différents compartiments racinaires (racines fines, souche et pivot, …) (Achat et al. 2018).
Une mortalité naturelle stochastique fixe ou variable (insectes, pathogènes) peut être prescrite. Il s‘y ajoute une mortalité climatique qui affecte les individus dont le bilan annuel en carbone est négatif, c’est à dire ceux dont le besoin en respiration de maintenance n’est pas couvert par l’assimilation photosynthétique. Cette mortalité climatique touche les individus de plus faible dimension durant les années à fort déficit hydrique.
8. Carbone du sol.
La dynamique du carbone du sol dérive du modèle Roth-C v6.3 (Coleman et Jenkinson, 1996) qui est appliqué ici en traitant l’ensemble du carbone du sol comme un ensemble de cinq pools et en conservant les valeurs des paramètres proposées par ses auteurs. La minéralisation du carbone du sol dépend ainsi de facteurs externes (température et humidité du sol), de la décomposabilité des apports en matière organique et de la teneur en argile du sol. L’effet du travail du sol est simulé dans GO+ en accélérant la vitesse de minéralisation du carbone dans la fraction du sol affectée pour une durée de 12 semaines (cf. §11. Gestion sylvicole).
9. Minéralomasse et exportations d’éléments minéraux.
Les cycles biogéochimiques des éléments minéraux majeurs ne sont pas décrits de façon dynamique dans le modèle v 26.12. Mais un module permettant de quantifier 1) les minéralomasses et les exportations de nutriments (N, P, K, Ca, Mg) avec les récoltes en fonction de l’itinéraire technique (pour toutes les essences étudiées) et 2) d’évaluer les bilans entrées-sorties (pour les peuplements de Pin maritime uniquement) a été inclus dans cette version. Pour chaque essence étudiée, le module prend en compte les concentrations en nutriments dans les différents compartiments de l’arbre ainsi que l’évolution des concentrations en fonction du stade de développement (Achat et al. 2018). Ce module est inactivé dans les simulations effectuées pour Forêts-21.
10. Production et récolte
Le volume et la biomasse récoltables sur pied (découpe 7cm) sont calculés annuellement. Les fractions récoltées et exportées hors parcelle (tronc, souche, branche, feuillage) sont prescrites dans l’ITK choisi, le reste retournant au sol. Le sous-étage n’est pas récolté.
11. Gestion sylvicole.
Les itinéraires techniques (ITK) sont une série d’opérations qui affectent le peuplement principal, le sous-étage et le sol. Ces opérations sont prescrites selon les combinaisons voulues de techniques de régénération, dégagement, éclaircies et récoltes. Elles peuvent être déclenchées par différents critères :
- Age des arbres ;
- Valeurs seuil de variables dendrométriques comme le diamètre ou la hauteur moyenne des troncs ;
- Indice de compétition (RDI, Le Moguedec et Dhôte, 2012) ;
Accèder aux itinéraires sylvicoles de Forêts-21.
Peuplement principal.
Les valeurs initiales de densité et la circonférence individuelle de chaque arbre sont prescrites. Deux régimes de conduite sont décrits dans la version 27.0 de GO+, la futaie régulière et le taillis.
La conduite en futaie régulière est définie par :
- les critères de déclenchement de chaque éclaircie et de la coupe finale. Ces critères ont été définis par les experts en sylviculture de chacune de essences prises en compte pour le projet Forêts-21. Ils peuvent être l’âge des arbres, un indice de compétition (RDI), la circonférencemoyenne ou dominante, ou la hauteur moyenne ou dominante, ou encore toute combinaison de ces critères.
- le critère de sélection et le pourcentage d’arbres récoltés. Les éclaircies peuvent être effectuées par le haut, par le bas ou de façon aléatoire.
- les fractions récoltées et exportées hors parcelle (tronc sur écorce, souche, branches, feuillage, grosses racines)
Le régime de taillis est caractérisé par le critère de déclenchement de la coupe, la densité des souches, le nombre de brins sélectionnés par souche et le nombre de recépées successives.
Sous–étage.
La biomasse du sous-étage est détruite partiellement ou totalement à chaque éclaircie et coupe. Elle est aussi détruite partiellement ou totalement par les opérations de dégagement qui peuvent être prescrites avant ou après régénération. La fraction de biomasse détruite par ces opérations peut prendre des valeurs différentes pour les parties aériennes et racinaires. Sol. Les opérations forestières affectent le sol par les retours au sol de matière organique morte provenant de la destruction du sous étage ou les résidus de récolte et des parties non récoltées du peuplement principal. Les opérations de travail et préparation du sol affectent aussi les processus de transformation et minéralisation de la matière organique du sol. Le modèle calcule la fraction de carbone du sol affectée par les opérations forestières (labour, disquage, rouleau lourd…). Pour cette fraction, la constante de décomposition K et le facteur de partitionnement en CO2du modèle Roth-C sont modifiés comme suit:
- Le K des quatre compartiments du carbone du sol RPM, DPM, HUM et BIO sont multipliées par un facteur q = 3.
- Le facteur de partitionnement en CO2, initialement fixé à 0.8, est porté à une valeur maximale de 1.
- Ces deux paramètres reprennent leur valeur initiale en suivant une diminution exponentielle sur une durée de 277 jours (taux de décroissance = 0,0025 jour-1).
Ce paramétrage a été calibré avec des données de terrain publiées (Arrouays et al., 1994 ; Jolivet, 2000) comme le détaille la thèse de Moreaux (2012).
12. Références citées.
Achat, D. L., Martel, S., Picart, D., Moisy, C., Augusto, L., Bakker, M. R., and Loustau, D. (2018) Modelling the nutrient cost of biomass harvesting under different silvicultural and climate scenarios in production forests, For EcolManag, 429, 642-653.
Arrouays, D. and Pelissier, P. (1994) Changes in Carbon Storage in Temperate Humic Loamy Soils after Forest Clearing and Continuous Corn Cropping in France, Plant and Soil, 160, 215-223. Bellassen, V., le Maire, G., Guin, O., Dhote, J. F., Ciais, P., and Viovy, N. (2011) Modelling forest management within a global vegetation model-Part 2: Model validation from a tree to a continental scale, Ecol Model, 222, 57-75.
Ciais, P., Loustau, D., Bosc, A., Ogée, J., Dufrêne, E., François, C., Viovy, N., and Delage, F. (2010)How will the production of French forests respond to climate change? An integrated analysis from site to country scale. In: Forests, carbon cycle and climate change, Loustau, D. (Ed.), Quae, Paris.
Coleman K., and Jenkinson:, D.D.: RothC - 26.3 –(1996) A model for the turnover of carbon in soil. , in: Evaluation of soil organic matter models using existing, long-term datasets., edited by: Powlson D.S., Smith P., and J.U., S., NATO ASI Series I, Springer Verlag, Heidelberg, Germany, 237-246.
Davi, H., Dufrêne, E., Granier, A., Le Dantec, V., Barbaroux, C., François, C., and Bréda, N. (2005) Modelling carbon and water cycles in a beech forest, Ecol Model, 185, 387-405.
DePury, D. G. G. and Farquhar, G. D. (1997)Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models, Plant Cell Environ, 20, 537-557.
Desprez-Loustau, M. L., and Dupuis, F. (1994) Variation in the phenology of shoot elongation between geographic provenances of maritime pine (Pinuspinaster)-implications for the synchrony with the phenology of the twisting rust fungus, Melampsorapinitorqua, Ann Sci Forest, 51, 553-568.
Dufrêne, E., Davi, H., François, C., Le Maire, G., Le Dantec, V., and Granier, A. (2005) Modelling carbon and water cycles in a beech forest. Part I: Model description and uncertainty analysis on modelled NEE, Ecol Model, 185, 407-436. https://doi.org/10.1016/j.ecolmodel.2005.01.004
Farquhar, G. D., von Caemmerer, S., and Berry, J. (1980) A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species, Planta, 149, 78-90.
Gash, J. H. C. (1979) Analytical model of rainfall interception by forests, Q J Roy Meteor Soc, 105, 43-55, 10.1002/qj.49710544304.
Harrington, C. A., Gould, P. J., and St Clair, J. B. (2010)Modeling the effects of winter environment on dormancy release of Douglas-fir, Forest EcolManag, 259, 798-808, 10.1016/j.foreco.2009.06.018, 2010
Jolivet, C. (2000)Le carbone organique des sols des Landes de Gascogne variabilité spatiale et effets des pratiques sylvicoles et agricoles, Thèse de doctorat en Sciences biologiques fondamentales et appliquées, Université de Bourgogne, Dijon, 306p.
Jones, H.G.: Plants and microclimate (1992) A quantitative approach to environmental plant physiology, Cambridge University Press, Cambridge.
Le Moguedec, G. and Dhote, J. F. (2012) Fagacees: a tree-centered growth and yield model for sessile oak (Quercuspetraea L.) and common beech (Fagus sylvatica L.), Ann ForSci, 69, 257-269.
Kramer, K. (1994) Selecting a Model to Predict the Onset of Growth of Fagus-Sylvatica, Journal of Applied Ecology, 31, 172-181.
Landsberg, J. J. and Waring, R. H. (1997)A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning, For EcolManag, 95, 209-228.
Loustau, D., Bosc, A., Colin, A., Ogee, J., Davi, H., Francois, C., Dufrene, E., Deque, M., Cloppet, E., Arrouays, D., Le Bas, C., Saby, N., Pignard, G., Hamza, N., Granier, A., Breda, N., Ciais, P., Viovy, N., and Delage, F. (2005) Modeling climate change effects on the potential production of French plains forests at the sub-regional level, Tree Physiol, 25, 813-823. https://doi.org/10.1093/treephys/25.7.813
Mohren, G. M. J., and Bartelink, H. H. (1990)Modeling the effects of needle mortality-rate and needle area distribution on dry-matter production of Douglas-fir,Neth J AgrSci, 38, 53-66.
Moreaux V. et al. (2020). Energy, water and carbon exchanges in managed forest ecosystems: description, sensitivity analysis and evaluation of the Inra GO+ model, version 27.0. Geoscientific Model Development.**(under review)**
Moreaux, V. (2012) Observation et modélisation des échanges d'énergie et de masse de jeunes peuplements forestiers du Sud-Ouest de la France., Ph. D.Thesis., Ecole Doctorale 304 "Sciences et Environnements", Thématique "Physique de l'Environnement", Université de Bordeaux-1, Bordeaux, 262 pages pp., 2012. https://www.theses.fr/2012BOR14506
Nakai, T., Sumida, A., Daikoku, K. i., Matsumoto, K., van der Molen, M. K., Kodama, Y., Kononov, A. V., Maximov, T. C., Dolman, A. J., Yabuki, H., Hara, T., and Ohta, T. (2008) Parameterisation of aerodynamic roughness over boreal, cool- and warm-temperate forests, Agr Forest Meteorol, 148, 1916-1925, https://doi.org/10.1016/j.agrformet.2008.03.009, 2008.
Ryan, M. G. (1991) Effects of climate change on plant respiration, EcolAppl, 1, 157-167, 1991.
Shaiek, O., Loustau, D., Trichet, P., Meredieu, C., Bachtobji, B., Garchi, S., and El Aouni, M. H. (2011) Generalized biomass equations for the main above-ground biomass components of maritime pine across contrasting environments, Ann Forest Sci, 68, 443-452.
Spitters, C. J. T., Toussaint, H. A. J. M., and Goudriaan, J. (1986) Separating the diffuse and direct component of global radiation and its implications for modeling canopy photosynthesis Part I. components of incoming radiation, Agric For Meteorol, 38, 217-229.
Wutzler, T., Wirth, C., and Schumacher, J. (2008) Generic biomass functions for Common beech (Fagus sylvatica) in Central Europe: predictions and components of uncertainty, Can J Forest Res, 38, 1661-1675, 10.1139/x07-194. https://www.nrcresearchpress.com/doi/full/10.1139/X07-194